
Prediction of the mechanism of Dictamni Cortex in treating dermatitis, eczema and psoriasis based on "the same treatment for different diseases"
-
摘要:目的
通过网络药理学和分子对接技术分析白鲜皮“异病同治”皮炎、湿疹、银屑病的分子靶点及其相关作用机理。
方法运用中药系统药理学数据库与分析平台(TCMSP)检索并筛选白鲜皮的主要活性成分, 并运用STP数据库预测有效成分的作用靶点。借助TTD、OMIM、DrugBank、GeneCards、DisGeNET数据库获取治疗皮炎、湿疹、银屑病的有关靶点,借助Venny软件获取皮炎、湿疹、银屑病的相同靶点以及与白鲜皮有效成分作用靶点的交集靶点。利用Cytoscape绘制网络图, STRING数据库构建白鲜皮治疗皮炎、湿疹、银屑病的相同靶基因的蛋白互作网络图。运用Metascape数据库进行基因本体(GO)富集分析和京都基因与基因组百科全书(KEGG)通路富集分析,并运用R语言进行可视化处理。通过Discovery Studio 2019 Client、AutoDock Tools、PyMOL进行分子对接和可视化。
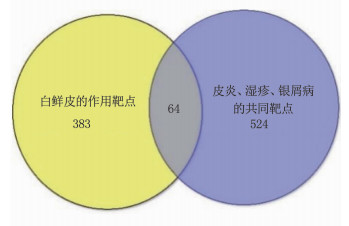
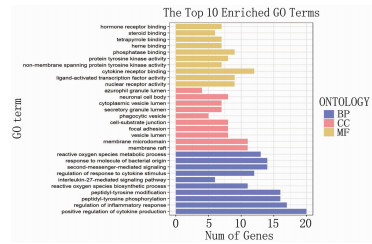
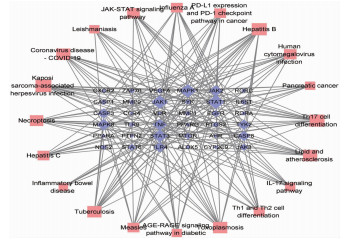
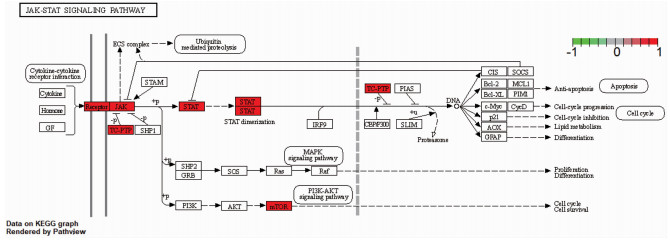
结果经筛选白鲜皮有效成分15个,与皮炎、湿疹、银屑病相同靶点的交集靶点共64个。白鲜皮活性成分治疗皮炎、湿疹、银屑病的核心靶点依次为肿瘤坏死因子(TNF)、Toll样受体4(TLR4)、血管内皮生长因子A (VEGFA)、信号转导和转录激活因子3(STAT3)、信号转导和转录激活因子1(STAT1)等。核心通路为高级糖基化终末产物-受体(AGE-RAGE)、白细胞介素17(IL-17)、酪氨酸激酶-信号传导及转录活化因子(JAK-STAT)信号通路等。
结论白鲜皮可能通过IL-17、JAK-STAT、AGE-RAGE等多个通路作用于TNF、TLR4、VEGFA等多个靶点,从而在皮炎、湿疹、银屑病中起到抗炎止痒的作用。
Abstract:ObjectiveTo analyze the molecular targets and related mechanisms of dermatitis, eczema, and psoriasis by Dictamni Cortex through network pharmacology and molecular docking based on "the same treatment for different diseases".
MethodsThe main active ingredients of Dictamni Cortex were searched and screened by using the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform(TCMSP) database, and the targets of active ingredients were predicted by the Swiss Target Prediction(STP) database. The relevant targets for the treatment of dermatitis, eczema, and psoriasis were obtained by TTD, OMIM, DrugBank, GeneCards, and DisGeNET database, and Venny software was used to obtain the same targets for dermatitis, eczema, and psoriasis, as well as targets that interacted with the active ingredients of Dictamni Cortex. Cytoscape was used to draw the network map, and the STRING database was used to construct the protein interaction network map of the same target genes of Dictamni Cortex in the treatment of dermatitis, eczema, and psoriasis. Metascape database was used for Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis, and R language was used for visualization. Molecular docking and visualization were performed through Discovery Studio 2019 Client, AutoDock Tools, and PyMOL.
ResultsA total of 15 active ingredients from Dictamni Cortex were obtained, and there were 64 intersection targets for the same targets of Dictamni Cortex and dermatitis, eczema, and psoriasis. The core targets of the active ingredients of Dictamni Cortex in the treatment of dermatitis, eczema, and psoriasis were tumor necrosis factor (TNF), toll-like receptor 4(TLR4), vascular endothelial growth factor A(VEGFA), signal transduction and transcription activator 3(STAT3), signal transduction and transcription activator 1(STAT1), etc. The core pathways included advanced glycosylation end-product-receptor (AGE-RAGE), interleukin-17 (IL-17), Janus kinase-signal transducer and activator of transcription (JAK-STAT) pathway, etc.
ConclusionDictamni Cortex may act on multiple targets such as TNF, TLR4 and VEGFA through multiple pathways including IL-17, JAK-STAT and AGE-RAGE to exert anti-inflammatory and anti-itch effects in dermatitis, eczema and psoriasis.
-
Keywords:
- Dictamni Cortex /
- dermatitis /
- eczema /
- psoriasis /
- network pharmacology /
- molecular docking /
- intersection target /
- signal pathway
-
脑血管病(CVD)是由多种脑血液循环障碍引起的脑部疾病的总称,具有高发病率、高病死率等特点[1]。全球疾病负担工作组(GBDWG)报告[2]显示,中国居民终生罹患CVD的风险高达39.3%, 远高于全球平均水平(24.9%)。尿酸(UA)是嘌呤代谢的终产物,具有抗氧化和清除氧自由基的作用[3]。研究[4]发现,血清尿酸(SUA)水平升高与CVD的发病率及病死率有关。荟萃分析[5-6]表明, SUA水平升高与中风、缺血性卒中(IS)发病风险增加有关; 但也有研究[7]提出高SUA水平可能对IS具有保护作用。目前, SUA在认知功能障碍方面的病理生理作用和临床意义仍存在争议[8]。一项长期队列研究[9]提出SUA水平升高可能显著增加罹患血管性痴呆(VaD)的风险; 但也有研究[10]发现SUA与VaD风险无相关性。
孟德尔随机化(MR)分析通过使用与暴露因素强相关性的单核苷酸多态性(SNPs)作为工具变量(IVs), 来推断暴露与结局的因果关系[11-12]。MR分析的优势在于能克服传统研究中混杂因素和反向因果关系的影响,从而得到更为可靠的结论[13-14]。本研究基于全基因组关联研究(GWAS)的汇总数据,应用MR分析探讨SUA与多种CVD的关联性[15], 包括中风, IS及其亚型如大动脉粥样硬化性卒中(LAS)、小血管性卒中(SVS)、心源性栓塞性卒中(CES), 以及VaD及其亚型如皮质下血管性痴呆(SIVaD)、多发梗死性痴呆(MID)、混合性痴呆(MixD)和其他原因性痴呆,以揭示SUA与这些疾病的双向因果关系,为CVD的预防和治疗提供新的科学依据。
1. 材料与方法
1.1 研究设计
本研究遵循孟德尔随机化研究报告规范(STROBE-MR)[14], 为了评估SUA与CVD的双向因果关系,分别进行了20次两样本MR分析。研究设计严格遵循MR研究的3个主要假设[16-18]: ①关联性假设,作为IVs的SNPs必须与暴露因素显著相关; ②独立性假设, IVs和混杂因素之间不存在相关性; ③排他性假设, IVs只能通过暴露与结局相关。本研究使用的数据均可公开获取,无需伦理批准和知情同意。研究设计见图 1。
![]() 图 1 孟德尔随机化研究设计图IVs: 工具变量; exposure: 暴露; outcome: 结局; confounders: 混杂因素; MR: 孟德尔随机化; serum uric acid levels: 血清尿酸水平; stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; other: 其他原因性痴呆。
图 1 孟德尔随机化研究设计图IVs: 工具变量; exposure: 暴露; outcome: 结局; confounders: 混杂因素; MR: 孟德尔随机化; serum uric acid levels: 血清尿酸水平; stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; other: 其他原因性痴呆。1.2 数据来源
本研究暴露和结局GWAS数据均来自于GWAS数据库(https://gwas.mrcieu.ac.uk/), 在该数据库检索框中分别输入“serum uric acid levels”“stroke”“ischemic stroke”“vascular dementia”等术语。所有研究对象均限定为欧洲血统,以尽量减少与种族因素相关的偏差。最终确定暴露因素SUA的GWAS数据由SAKAUE S等[19](PMID: 34594039)发表。结局中风、IS及其亚型相关的GWAS数据由MALIK R等[20](PMID: 29531354)发表。VaD及其亚型的GWAS数据来自FinnGen。见表 1。
表 1 本研究使用GWAS数据集的详细信息表型 数据来源 研究人群 样本量/例 单核苷酸多态性/个 PMID 血清尿酸水平 SAKAUE S等 欧洲人群 343 836 19 041 286 34594039 中风 MALIK R等 欧洲人群 446 696 8 211 693 29531354 缺血性卒中 MALIK R等 欧洲人群 440 328 8 296 492 29531354 大动脉粥样硬化性卒中 MALIK R等 欧洲人群 410 484 8 418 349 29531354 小血管性卒中 MALIK R等 欧洲人群 198 048 8 280 845 29531354 心源性栓塞性卒中 MALIK R等 欧洲人群 413 304 7 954 834 29531354 血管性痴呆 FinnGen 欧洲人群 212 389 16 380 457 — 皮质下血管性痴呆 FinnGen 欧洲人群 211 554 16 380 455 — 多发梗死性痴呆 FinnGen 欧洲人群 211 494 16 380 454 — 混合性痴呆 FinnGen 欧洲人群 211 398 16 380 453 — 其他原因性痴呆 FinnGen 欧洲人群 211 687 16 380 454 — 1.3 工具变量筛选
根据STROBE-MR[17], 本研究的SNPs采取以下步骤进行筛选: ①以P<5×10-8为阈值筛选出与暴露强相关的SNPs作为IVs。②设置连锁不平衡系数r2=0.001、区域宽度为10 000 kb以确保所选的SNPs位点相互独立[21]。③提取与结局相关的SNPs, 整合暴露和结局数据集,并删除回文序列。④通过PhenoScanner数据库(http://www.phenoscanner.medschl.cam.ac.uk/)排除混杂因素。⑤计算每个IVs的R2和F统计量。剔除F值< 10的弱IVs, 确保所选IVs的有效性[22]。⑥采用MR-PRESSO识别异常值和校正水平多效性。反向MR分析,从不同CVD的GWAS数据中筛选出独立的SNPs作为IVs(P < 5×10-6), 其他筛选标准与正向MR分析一致。
1.4 统计学分析
本研究主要采用逆方差加权法(IVW), 以加权中位数法(weighted median)、加权模式方法(weighted mode)和MR-Egger回归法作为补充,评估SUA与CVD的因果关联[23], 以森林图呈现分析结果。采用Cochran′s Q检验判断SNPs间的异质性[24-25]。为了识别和减少潜在的水平多效性引起的偏差,本研究采用MR-PRESSO[26]和MR-Egger[27]截距法进行检测。通过“留一法”敏感性分析,依次剔除单个SNP, 以评估其是否对整体结果有影响。通过观察散点图和漏斗图的对称性来检查结果是否稳定。所有分析均在R软件(4.3.2版)中进行,使用“TwoSampleMR(0.5.9版)”和“MR-PRESSO(1.0版)”[28]这2个R包。统计学显著性通过Bonferroni校正后确定, P < 0.005被认为具有显著性, P≥0.005~0.05表示潜在的因果关联, P>0.05为差异无统计学意义。
2. 结果
2.1 SUA水平与不同CVD因果关联的正向MR分析结果
2.1.1 IVs的选择结果
根据上述IVs的筛选标准及步骤,本研究选择了不同数量的SNPs作为评估SUA水平与不同CVD因果关联的IVs, 筛选出163、164、169、168、170、165、171、179、180和181个SNPs分别作为中风、IS、LAS、SVS、CES、VaD、SIVaD、MID、MixD和其他原因性痴呆的IVs。
2.1.2 SUA水平与中风、IS及其亚型之间的因果关联
IVW分析结果显示,基因预测的SUA水平与中风(OR=1.183, 95% CI: 1.081~1.295, P=2.51×10-4)、IS(OR=1.196, 95% CI: 1.084~1.320, P=3.81×10-4)和LAS(OR=1.466, 95% CI: 1.134~1.897, P=0.004)的发病风险呈正相关。上述结果经Bonferroni校正P < 0.005后,差异仍有统计学意义。基因预测的SUA水平与SVS(OR=1.159, 95% CI: 0.917~1.467, P=0.218)、CES (OR=0.878, 95% CI: 0.743~1.037, P=0.126)之间不存在因果关联。
2.1.3 SUA水平与VaD及其亚型之间的因果关联
IVW分析结果显示,基因预测的SUA水平与VaD(OR=0.451, 95% CI: 0.273~0.745, P=0.002)和MID(OR=0.372, 95% CI: 0.144~0.959, P=0.041)的发病风险呈负相关。上述结果经Bonferroni校正P < 0.005后,提示SUA与MID存在潜在的因果关联。基因预测的SUA水平与SIVaD(OR=0.859, 95% CI: 0.371~1.986, P=0.722)、MixD(OR=1.693, 95% CI: 0.444~6.462, P=0.441)和其他原因性痴呆(OR=1.104, 95% CI: 0.545~2.237, P=0.784)之间不存在因果关联。
2.1.4 敏感性分析
Cochran′s Q检验结果显示,上述结果均不存在异质性(P中风=0.845, PIS=0.965, PLAS=0.189, PSVS=0.984, PCES=0.957, PVaD=0.855, PSIVaD=0.930, PMID=0.538, PMixD=0.999,P其他原因性痴呆=0.194)。MR-Egger截距检测结果也显示均不存在水平多效性(P中风=0.360, PIS=0.354, PLAS=0.786, PSVS=0.858, PCES=0.331, PVaD=0.772, PSIVaD=0.659, PMID=0.421, PMixD=0.775, P其他原因性痴呆=0.279)。此外, MR-PRESSO分析在SUA水平与LAS风险的因果关联中识别出4个异常SNPs(rs146787580、rs7696556、rs34868798、rs9807214), 在对这些SNPs进行校正后, SUA水平对LAS风险的因果效应依然显著,其余均未检测出异常SNPs。留一法分析表明消除任何一个SNP均不会对整体结果有影响。最后, SUA水平对不同CVD风险预测的散点图和漏斗图显示了因果效应的分布是对称的,这进一步验证了结果的稳定性和可靠性。SUA水平与不同CVD因果关联的正向MR分析结果见图 2。
![]() 图 2 正向孟德尔随机化分析结果IVW: 逆方差加权法; Stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; Dementia of other causes: 其他原因性痴呆。
图 2 正向孟德尔随机化分析结果IVW: 逆方差加权法; Stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; Dementia of other causes: 其他原因性痴呆。2.2 SUA水平与不同CVD因果关联的反向MR分析结果
2.2.1 IVs的选择结果
本研究选择了不同数量的SNPs作为评估不同CVD与SUA水平因果关联的IVs, 筛选出中风(35个)、IS(34个)、LAS(25个)、SVS(23个)、CES(18个)、VaD(6个)、SIVaD(12个)、MID(7个)、MixD(3个)、其他原因性痴呆(4个)分别作为SUA的IVs。
2.2.2 不同CVD与SUA水平之间的因果关联
IVW分析结果显示,基因预测的中风(OR=1.007, 95% CI: 0.994~1.021, P=0.262)、IS(OR=1.004, 95% CI: 0.991~1.016, P=0.572)、LAS(OR=1.005, 95% CI: 0.999~1.010, P=0.086)、SVS(OR=0.995, 95% CI: 0.988~1.001, P=0.101)、CES(OR=1.011, 95% CI: 1.000~1.023, P=0.058)、VaD(OR=1.002, 95% CI: 0.999~1.005, P=0.278)、SIVaD(OR=1.001, 95% CI: 0.999~1.003, P=0.273)、MID(OR=1.003, 95% CI: 0.999~1.006, P=0.103)、MixD(OR=1.003, 95% CI: 0.999~1.008, P=0.157)、其他原因性痴呆(OR=1.004, 95% CI: 0.999~1.010, P=0.135)与SUA水平之间均不存在因果关联。
2.2.3 敏感性分析
Cochran′s Q检验结果显示均不存在异质性(P中风=0.384, PIS=0.265, PLAS=0.611, PSVS=0.971, PCES=0.087, PVaD=0.727, PSIVaD=0.329, PMID=0.299, PMixD=0.165, P其他原因性痴呆=0.240)。MR-Egger截距检测结果也显示均不存在水平多效性(P中风=0.813, PIS=0.661, PLAS=0.224, PSVS=0.439, PCES=0.919, PVaD=0.218, PSIVaD=0.060, PMID=0.654, PMixD=0.363, P其他原因性痴呆=0.220)。此外, MR-PRESSO对异常值进行检测,在中风与SUA的因果关联中识别出2个异常SNPs(rs147076266、rs10774624), 在IS与SUA的因果关联中识别出2个异常SNPs(rs7099238、rs60102266), 在LAS与SUA的因果关联中识别出1个异常SNP(rs142532801), 在SVS与SUA的因果关联中识别出1个异常SNP(rs75501811), 在CES与SUA的因果关联中识别出1个异常SNP(rs13143308), 但对异常值进行校正后,所有因果效应依然显著,其余均未检测出异常SNPs。留一法分析结果表明MR分析结果稳健。最后,不同CVD风险对SUA水平预测的散点图和漏斗图显示了因果效应的分布是对称的,提示了结果的可靠性。SUA水平与不同CVD因果关联的反向MR分析结果见图 3。
![]() 图 3 反向孟德尔随机化分析结果IVW: 逆方差加权法; Stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; Dementia of other causes: 其他原因性痴呆。
图 3 反向孟德尔随机化分析结果IVW: 逆方差加权法; Stroke: 中风; IS: 缺血性卒中; LAS: 大动脉粥样硬化性卒中; SVS: 小血管闭塞性卒中; CES: 心源性栓塞型卒中; VaD: 血管性痴呆; SIVaD: 皮质下血管性痴呆; MID: 多发梗死性痴呆; MixD: 混合性痴呆; Dementia of other causes: 其他原因性痴呆。3. 讨论
本研究通过双样本双向MR分析,深入探讨了SUA水平与不同CVD之间的因果关联。正向MR分析结果显示, SUA水平升高会增加中风、IS和LAS的发病风险,但也会降低VaD、MID的发病风险。然而, MR分析并未发现SUA与SVS、CES、SIVaD、MixD和其他原因性痴呆之间存在因果关系。此外,反向MR分析结果并不支持基因预测的CVD风险对SUA水平存在因果影响。这些发现为了解SUA在CVD中的作用提供了新的视角,有助于开发针对性的预防和治疗措施,以降低特定患者群体的疾病风险。
既往大量观察性研究分析了SUA与CVD的相关性。LI M等[29]进行了一项回顾性分析发现,高SUA水平与中风发病风险增加相关(RR=1.22, 95% CI: 1.02~1.46), 即使在调整了年龄、高血压、糖尿病和胆固醇等已知风险因素后,该相关性依然显著。WANG C Q等[30]通过中介MR研究也发现, SUA水平升高与中风、IS、LAS风险增加呈正相关。本研究使用不同来源的GWAS数据进行MR分析,得出了类似的结论,即SUA每增加1个单位,中风、IS、LAS的发病风险分别增加18.3%、19.6%、46.6%。考虑到本研究MR分析提取了更多符合条件的IVs,并且仅使用了来自欧洲血统的GWAS汇总数据,这使得本研究结果更加准确。
作为一个严重的公共健康问题, VaD的发病率呈上升趋势[31]。多个观察性研究探讨了SUA水平与VaD风险之间的相关性。一项基于人群的队列研究[32]发现,SUA水平升高与VaD风险降低有关,并指出其具有神经保护作用。XU Y Z等[33]研究也指出,较低的SUA水平与认知功能下降有关。然而,一项前瞻性队列研究[34]显示,当人体SUA水平升高>400 μmol/L时, VaD的风险可能会增加50%(HR=1.50, 95% CI: 1.28~1.77)。本研究结果与部分既往研究结果并不完全一致,其原因可能为: ①既往研究主要是观察性研究,结果可能受到混杂因素和反向因果关系的影响。②部分研究可能因样本量不足而限制了结论的普遍性和准确性。
SUA对CVD的作用机制可能涉及多种生物化学过程和炎症反应。有研究[35]提出SUA可能通过抑制一氧化氮(NO)合酶的活性,间接促进高血脂和高血压的发展,这可能导致动脉壁上脂质的积累而形成动脉粥样硬化斑块,从而加剧CVD的发生。有研究[36]指出在由低密度脂蛋白(LDL)引发的炎症反应中,SUA可能作为助氧化剂,降低NO水平,导致血管损伤。此外, SUA还可能通过增强炎症因子的活性,加剧血管内皮的炎症反应,促进血管内皮损伤,增加动脉粥样硬化的风险,这也是CVD的一个主要病因[37-38]。上述研究表明,SUA可能通过激活炎症途径和促进氧化应激参与机体的炎症损伤过程,破坏血管内皮细胞,促进动脉粥样硬化的发展。同时,炎症因子可通过影响黄嘌呤氧化酶的活性,不仅增加UA的生成,还可能导致细胞损伤或凋亡[39],形成UA与炎症之间的恶性循环,从而可能增加CVD的风险。临床上对于SUA水平较高的患者,应积极采取预防措施以降低CVD的发病风险。
本研究的反向MR分析结果未发现CVD风险与SUA水平之间存在因果关联,这一发现与部分观察性研究结果不同。既往研究[40]指出,随着IS发作后时间的推移,SUA水平呈上升趋势,且与患者长期不良预后相关。然而,也有研究[41]显示在脑血管事件发生后SUA的水平显著下降,这意味着SUA可能参与了疾病的进展过程。UA具有抗氧化性质,在大脑缺血、缺氧的情况下,脑内脂质过氧化物增加以及组织抗氧化水平降低都是氧化应激增加的间接证据。
本研究的优势: ①利用遗传学方法证明SUA水平与CVD风险之间的双向因果关联,有效避免了传统观察性研究中的混杂因素和反向因果关系的影响。②通过使用大规模GWAS的汇总数据并结合敏感性分析,进一步证实了本研究结果的可靠性。本研究的局限性: ①研究依赖于GWAS的汇总数据,缺乏个体层面的数据,这限制了进行年龄、性别等特定亚组分析的能力,以及评估不同亚组间因果效应差异的可能性。②所使用的GWAS数据主要来源于欧洲人群,因此研究结果在其他种族或人群的普适性需要进一步验证。
综上所述,本研究MR分析表明SUA水平升高与中风、IS、LAS发病风险呈正相关,与VaD、MID发病风险呈负相关,为临床上采取严格控制SUA水平的干预措施来降低CVD的发病风险提供科学依据。未来可通过扩大样本量范围,涵盖多样化人群,以探讨特定的亚组或不同人群中可能存在的因果关联,进一步开发更为精准的预防和治疗策略。
-
![]()
图 10 前茵芋碱与TNF、白鲜明碱与EGFR、赤霉素B与MMP9、赤霉素B与PPARG的分子结合模式
A: 前茵芋碱与TNF的分子结合; B: 白鲜明碱与EGFR的分子结合; C: 赤霉素B与MMP9的分子结合; D: 赤霉素B与PPARG的分子结合。
表 1 白鲜皮的有效成分
序号 MOL ID 成分名称 DL/% OB 1 MOL006271 赤霉素B 0.27 35.51 2 MOL000098 槲皮素 0.28 46.43 3 MOL006268 前茵芋碱 0.21 42.14 4 MOL000006 木犀草素 0.25 36.16 5 MOL006235 白鲜明碱 0.21 59.14 6 MOL000173 汉黄芩素 0.23 30.68 7 MOL002663 茵芋碱 0.2 40.14 8 MOL013352 黄柏酮 0.77 43.29 9 MOL006261 异斑点沸林草碱 0.2 31.99 10 MOL005043 菜油甾醇 0.71 37.58 11 MOL000358 β-谷甾醇 0.75 36.91 12 MOL006272 9α-羟基梣酮-9-O-β-D-葡萄糖苷 0.62 55.52 13 MOL006262 7α-乙酰叶枯醇 0.71 36.08 14 MOL006233 3′-O-甲基花旗松素 0.3 48.36 15 MOL006270 甘遂烷 0.75 44.02 PL: 药物相似性; OB: 药物生物口服利用度。  下载: 导出CSV
下载: 导出CSV
表 2 关键化合物与核心蛋白结合能
对接化合物 结合能量/(kcal/mol) TNF EGFR MMP9 PPARG 赤霉素B -8.13 -6.47 -7.82 -7.48 甘遂烷 -8.48 -6.33 -7.61 -6.66 前茵芋碱 -8.49 -5.72 -5.72 -6.21 7α-乙酰叶枯醇 -7.87 -5.26 -6.48 -5.95 白鲜明碱 -6.01 -6.88 -5.84 -5.98
下载: 导出CSV
表 3 关键化合物与核心靶点氢键结合数量与位点
化合物与靶点 氢键数量 结合位点 前茵芋碱与TNF 2 ARG-107、ARG-108 白鲜明碱与EGFR 3 ASN-771、ASN-826、ARG-831 赤霉素B与MMP9 1 PHE-250 赤霉素B与PPARG 1 ASN-412
下载: 导出CSV
-
[1] ELSNER P, AGNER T. Hand eczema: treatment[J]. J Eur Acad Dermatol Venereol, 2020, 34(Suppl 1): 13-21.
[2] CHEN Y C, YAN Y H, LIU H Z, et al. Dihydroartemisinin ameliorates psoriatic skin inflammation and its relapse by diminishing CD8+ T-cell memory in wild-type and humanized mice[J]. Theranostics, 2020, 10(23): 10466-10482. doi: 10.7150/thno.45211
[3] 陆家睛, 史玉玲. 糖皮质激素在皮肤科的应用[J]. 皮肤科学通报, 2021, 38(4): 304-311. https://www.cnki.com.cn/Article/CJFDTOTAL-ZYXW202104004.htm [4] TOKUYAMA M, MABUCHI T. New treatment addressing the pathogenesis of psoriasis[J]. Int J Mol Sci, 2020, 21(20): 7488. doi: 10.3390/ijms21207488
[5] 王启斌, 袁昊, 马朝晖, 等. 中药液体制剂在炎性皮肤病中的应用[J]. 江西中医药, 2021, 52(10): 73-76. https://www.cnki.com.cn/Article/CJFDTOTAL-JXZY202110035.htm [6] GAO P, WANG L, ZHAO L, et al. Rapid identification, isolation, and evaluation on anti-neuroinflammatory activity of limonoids derivatives from the root bark of Dictamnus dasycarpus[J]. J Pharm Biomed Anal, 2021, 200: 114079. doi: 10.1016/j.jpba.2021.114079
[7] 陈佳骏, 杨妞妞. 白鲜皮及其活性成分抗炎止痒机制研究进展[J]. 江西中医药, 2020, 51(5): 77-80. https://www.cnki.com.cn/Article/CJFDTOTAL-JXZY202005033.htm [8] 邵海峰, 杨妞妞, 刘延庆. 基于数据挖掘的中医治疗湿疹用药规律分析[J]. 实用临床医药杂志, 2021, 25(24): 83-86, 96. doi: 10.7619/jcmp.20212927 [9] YANG S C, SUN F, RUAN J Y, et al. Anti-inflammatory constituents from cortex dictamni[J]. Fitoterapia, 2019, 134: 465-473. doi: 10.1016/j.fitote.2019.03.026
[10] ZHOU Z C, CHEN B, CHEN S M, et al. Applications of network pharmacology in traditional Chinese medicine research[J]. Evid Based Complement Alternat Med, 2020, 2020: 1646905.
[11] RU J L, LI P, WANG J N, et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines[J]. J Cheminform, 2014, 6: 13. doi: 10.1186/1758-2946-6-13
[12] DONG Y Z, ZHAO Q L, WANG Y G. Network pharmacology-based investigation of potential targets of astragalus membranaceous-Angelica sinensis compound acting on diabetic nephropathy[J]. Sci Rep, 2021, 11(1): 19496. doi: 10.1038/s41598-021-98925-6
[13] 胡伟, 李晨, 王晨光, 等. 基于UPLC-Q-Qrbitrap HRMS和网络药理学探究芪胶升白胶囊治疗白细胞减少症的作用机制[J]. 现代药物与临床, 2023, 9(2): 289-299. https://www.cnki.com.cn/Article/CJFDTOTAL-GWZW202302006.htm [14] UNIPROT CONSORTIUM. UniProt: a worldwide hub of protein knowledge[J]. Nucleic Acids Res, 2019, 47(D1): D506-D515. doi: 10.1093/nar/gky1049
[15] ZHOU Y Y, ZHOU B, PACHE L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets[J]. Nat Commun, 2019, 10(1): 1523. doi: 10.1038/s41467-019-09234-6
[16] YANG X H, LIU H H, LIU J N, et al. Rational selection of the 3D structure of biomacromolecules for molecular docking studies on the mechanism of endocrine disruptor action[J]. Chem Res Toxicol, 2016, 29(9): 1565-1570. doi: 10.1021/acs.chemrestox.6b00245
[17] ZHANG B Y, ZHAO J, WANG Z, et al. Identification of multi-target anti-AD chemical constituents from traditional Chinese medicine formulae by integrating virtual screening and in vitro validation[J]. Front Pharmacol, 2021, 12: 709607. doi: 10.3389/fphar.2021.709607
[18] LI B J, RUI J Q, DING X J, et al. Exploring the multicomponent synergy mechanism of Banxia Xiexin Decoction on irritable bowel syndrome by a systems pharmacology strategy[J]. J Ethnopharmacol, 2019, 233: 158-168. doi: 10.1016/j.jep.2018.12.033
[19] KRIEF S, MARTIN M T, GRELLIER P, et al. Novel antimalarial compounds isolated in a survey of self-medicative behavior of wild chimpanzees in Uganda[J]. Antimicrob Agents Chemother, 2004, 48(8): 3196-3199. doi: 10.1128/AAC.48.8.3196-3199.2004
[20] MANNINO G, IOVINO P, LAURIA A, et al. Bioactive triterpenes of Protium heptaphyllum gum resin extract display cholesterol-lowering potential[J]. Int J Mol Sci, 2021, 22(5): 2664. doi: 10.3390/ijms22052664
[21] YOON J Y, WANG J Y, ROEHRL M H A. An investigation into the prognostic significance of high proteasome PSB7 protein expression in colorectal cancer[J]. Front Med, 2020, 7: 401. doi: 10.3389/fmed.2020.00401
[22] PŁÓCIENNIKOWSKA A, HROMADA-JUDYCKA A, BORZECKA K, et al. Co-operation of TLR4 and raft proteins in LPS-induced pro-inflammatory signaling[J]. Cell Mol Life Sci, 2015, 72(3): 557-581. doi: 10.1007/s00018-014-1762-5
[23] 邓斌, 罗庆, 宋关斌. COX2通过HIF-1α/VEGFA/PDGFB信号通路增强炎性肌腱细胞的促血管新生能力[J]. 医用生物力学, 2021, 36(S01): 188-188. https://www.cnki.com.cn/Article/CJFDTOTAL-YISX2021S1359.htm [24] BENHADOU F, GLITZNER E, BRISEBARRE A, et al. Epidermal autonomous VEGFA/Flt1/Nrp1 functions mediate psoriasis-like disease[J]. Sci Adv, 2020, 6(2): eaax5849. doi: 10.1126/sciadv.aax5849
[25] BUTTURINI E, CARCERERI DE PRATI A, MARIOTTO S. Redox regulation of STAT1 and STAT3 signaling[J]. Int J Mol Sci, 2020, 21(19): 7034. doi: 10.3390/ijms21197034
[26] JU X, YANG X, YAN T, et al. EGFR inhibitor, AG1478, inhibits inflammatory infiltration and angiogenesis in mice with diabetic retinopathy[J]. Clin Exp Pharmacol Physiol, 2019, 46(1): 75-85. doi: 10.1111/1440-1681.13029
[27] KIM E N, KAYGUSUZ O, LEE H S, et al. Simultaneous quantitative analysis of ginsenosides isolated from the fruit of Panax ginseng C. A. Meyer and regulation of HO-1 expression through EGFR signaling has anti-inflammatory and osteogenic induction effects in HPDL cells[J]. Molecules, 2021, 26(7): 2092. doi: 10.3390/molecules26072092
[28] AL BARASHDI M A, ALI A, MCMULLIN M F, et al. Protein tyrosine phosphatase receptor type C (PTPRC or CD45)[J]. J Clin Pathol, 2021, 74(9): 548-552. doi: 10.1136/jclinpath-2020-206927
[29] SU W X, ZHAO Y, WEI Y Q, et al. Exploring the pathogenesis of psoriasis complicated with atherosclerosis via microarray data analysis[J]. Front Immunol, 2021, 12: 667690. doi: 10.3389/fimmu.2021.667690
[30] LIU C, TATE T, BATOURINA E, et al. Pparg promotes differentiation and regulates mitochondrial gene expression in bladder epithelial cells[J]. Nat Commun, 2019, 10(1): 4589.
[31] RUMZHUM N N, AMMIT A J. Cyclooxygenase 2: its regulation, role and impact in airway inflammation[J]. Clin Exp Allergy, 2016, 46(3): 397-410.








计量
- 文章访问数: 149
- HTML全文浏览量: 50
- PDF下载量: 14
 苏公网安备 32100302010246号
苏公网安备 32100302010246号
