
Study on total flavonoids from Selaginella uncinata in inhibiting malignant behavior of colorectal cancer cells through circular RNA circ_0006528 pathway
-
摘要:目的 探讨翠云草总黄酮对结直肠癌细胞恶性生物学行为的影响及可能机制。方法 体外培养结直肠癌细胞Caco-2, 用不同剂量(10、20、40 μg/mL)翠云草总黄酮干预24 h、或转染环状RNA circ_0006528小干扰RNA至Caco-2细胞后培养24 h、或用40 μg/mL的翠云草总黄酮干预转染circ_0006528过表达载体的Caco-2细胞24 h, 然后用检测细胞存活与生长比色法(MTT)、克隆形成实验、划痕实验和流式细胞术分别检测细胞活性、克隆形成数、迁移和凋亡。Western Blot检测Bax和Bcl-2蛋白表达,实时荧光定量聚合酶链反应(qRT-PCR)检测circ_0006528和miR-330-3p表达。双荧光素酶报告基因实验验证circ_0006528和miR-330-3p调控关系。结果 Caco-2细胞经翠云草总黄酮干预后,细胞活性、克隆形成数、迁移距离和细胞中Bcl-2蛋白、circ_0006528的表达均降低,而凋亡率和Bax蛋白、miR-330-3p的表达升高,差异有统计学意义(P < 0.05), 且呈剂量依赖性。干扰circ_0006528后, Caco-2细胞活性、克隆形成数、迁移距离和Bcl-2蛋白表达降低,细胞凋亡率和Bax蛋白表达升高,差异有统计学意义(P < 0.05)。circ_0006528靶向负调控miR-330-3p。过表达circ_0006528逆转了翠云草总黄酮对Caco-2细胞增殖、迁移和凋亡及miR-330-3p表达的影响。结论 翠云草总黄酮可能通过调控circ_0006528/miR-330-3p轴,抑制结直肠癌细胞增殖和迁移,并促进细胞凋亡。Abstract:Objective To investigate the effect of total flavonoids from Selaginella uncinata on malignant biological behavior of colorectal cancer cells and its possible mechanism.Methods Colorectal cancer cells Caco-2 were cultured in vitro. After Caco-2 cells were treated with different concentrations (10, 20 and 40 μg/mL) of total flavonoids from Selaginella uncinata for 24 hours, or Caco-2 cells were transfected with circ_0006528 small interfering RNA, or Caco-2 cells that transfected with circ_0006528 overexpression vector were treated with 40 μg/mL total flavonoids from Selaginella uncinata for 24 hours, methyl thiazolyl tetrazolium (MTT) assay, clone formation assay, scratch assay and flow cytometry were used to detect cell proliferation viability, the number of colony formation, migration and apoptosis, respectively. The expressions of Bax and Bcl-2 proteins were detected by western blot, and the expressions of circ_0006528 and miR-330-3p were detected by quantitative real-time polymerase chain reaction (qRT-PCR). The regulatory relationship between circ_0006528 and miR-330-3p was verified by dual luciferase reporter gene assay.Results After Caco-2 cells were intervened with total flavonoids from Selaginella uncinata, cell viability, the number of clone formation, migration distance and the expression of Bcl-2 protein as well as circ_0006528 in Caco-2 cells were significantly decreased, but apoptosis rate and the expression of Bax protein and miR-330-3p were significantly increased (P < 0.05), and they were dose-dependent. After circ_0006528 interference, Caco-2 cell activity, the number of clone formation, migration distance and Bcl-2 protein expression were significantly decreased, while apoptosis rate and Bax protein expression were significantly increased (P < 0.05). The circ_0006528 negatively regulated miR-330-3p by targeting. Overexpression of circ_0006528 reversed the effects of total flavonoids from Selaginella uncinata on proliferation, migration and apoptosis of Caco-2 cells and expression of miR-330-3p.Conclusion The total flavonoids from Selaginella uncinata may inhibit the proliferation and migration of colorectal cancer cells and promote cells apoptosis by regulating the circ_0006528/miR-330-3p axis.
-
阻塞性睡眠呼吸暂停低通气综合征(OSAHS)是一种以睡眠期间上呼吸道反复阻塞而导致气流中断或减少为特征的临床综合征,患者可表现为夜间打鼾、白天嗜睡、情绪障碍等,与心脑血管疾病、内分泌与代谢性疾病及神经认知功能障碍等密切相关[1]。近年来,随着OSAHS研究的不断深入, OSAHS伴发的2型糖尿病逐渐引起关注。OSAHS已被证实是2型糖尿病的独立危险因素之一,且在2型糖尿病的发展中发挥重要作用[2]。相关研究[3]表明, OSAHS患者的2型糖尿病患病率为15%~30%, 且重度OSAHS患者的患病率可能更高。反复呼吸阻塞或暂停引起的间歇性缺氧(IH)是低通气的特征性病理改变,最终导致OSAHS患者以低氧血症、高碳酸血症为主的病理生理基础[4]。研究[2]证实, IH通过多种机制参与2型糖尿病的发病过程。夜间脉搏血氧饱和度(SpO2)监测可反映睡眠期间血氧状态,已被应用于OSAHS和2型糖尿病等多种疾病的研究中,但关于OSAHS患者SpO2与发生2型糖尿病关系的研究目前尚少见。本研究探讨了OSAHS患者夜间SpO2水平与2型糖尿病之间的关系,并分析SpO2对OSAHS患者伴发2型糖尿病的预测价值,现报告如下。
1. 资料与方法
1.1 一般资料
选取2021年1—9月因打鼾至扬州大学附属医院门诊或住院申请行多导睡眠图仪(PSG)监测的患者作为研究对象。结合患者病史、体格检查及相关实验室检查结果,根据《阻塞性睡眠呼吸暂停低通气综合征诊治指南(草案)》[5]中OSAHS的诊断标准及2013年版《中国2型糖尿病防治指南》[6]中的糖尿病诊断标准,将纳入患者分为单纯OSAHS组76例和OSAHS伴2型糖尿病组77例。纳入标准: ①年龄18~80岁者; ②确诊OSAHS者。排除标准: ①中枢性睡眠呼吸暂停低通气综合征患者; ②发作性睡病患者; ③接受过OSAHS治疗者; ④妊娠期妇女; ⑤合并急性呼吸道感染者; ⑥严重肺部疾病如慢性阻塞性肺疾病、支气管哮喘、间质性肺疾病、慢性呼吸衰竭等患者; ⑦心功能不全者; ⑧合并糖尿病急性并发症者。
1.2 研究方法
1.2.1 临床资料收集
通过电子病历系统搜索并记录所有入组患者的性别、年龄、身高和体质量,计算体质量指数(BMI); 收集每例患者既往病史[高血压病、动脉粥样硬化性心血管疾病、高脂血症和糖尿病家族史(一级亲属患有2型糖尿病)]、个人史(吸烟史、饮酒史)等信息。
1.2.2 睡眠指标监测
在睡眠室患者信息系统根据住院号或门诊号搜索所有患者的睡眠指标结果并记录,包括呼吸暂停低通气指数(AHI)、氧减指数(ODI)、平均脉搏血氧饱和度(MSpO2)和最低脉搏血氧饱和度(LSpO2)。所有接受PSG监测的患者均在医院睡眠室进行整夜(至少7 h)睡眠监测,并监测鼾声、胸腹呼吸运动、口鼻气流和体位等。SpO2由指触式脉搏血氧饱和度仪监测获得,所有患者监测当天禁饮酒、咖啡,禁服镇静剂及催眠药。
1.3 统计学分析
采用SPSS 20.0统计学软件对数据进行处理,计量资料进行正态性和方差齐性检验,符合正态分布者以(x±s)表示, 2组比较行独立样本t检验,计数资料以[n(%)]表示, 2组比较行χ2检验。OSAHS患者伴2型糖尿病的影响因素分析采用二元Logistic回归分析。采用受试者工作特征(ROC)曲线分析夜间SpO2对OSAHS患者伴2型糖尿病的预测价值。以P<0.05为差异有统计学意义。
2. 结果
2.1 一般资料及睡眠监测指标比较
与单纯OSAHS组比较, OSAHS伴2型糖尿病组患者年龄较大, BMI、AHI、ODI水平和高脂血症患病率、有糖尿病家族史者占比较高, MSpO2、LSpO2水平较低,差异有统计学意义(P<0.05)。2组患者在性别、吸烟史、饮酒史、高血压病及动脉粥样硬化性心血管疾病患病率方面比较,差异无统计学意义(P>0.05)。见表 1。
表 1 2组患者一般资料与睡眠监测指标水平比较(x±s) [n(%)]指标 OSAHS伴2型糖尿病组(n=77) 单纯OSAHS组(n=76) t/χ2 P 年龄/岁 59.19±12.17 54.42±13.14 -2.332 0.021 性别 男 51(66.23) 56(73.68) 1.010 0.315 女 26(33.77) 20(26.32) BMI/(kg/m2) 27.65±4.20 25.52±2.85 -3.680 <0.001 吸烟史 26(33.77) 22(28.95) 0.413 0.521 饮酒史 24(31.17) 22(28.95) 0.09 0.764 高血压病 60(77.92) 55(72.37) 0.632 0.427 高脂血症 46(59.74) 28(36.84) 8.03 0.005 动脉粥样硬化性心血管疾病 31(40.26) 36(47.37) 0.785 0.376 糖尿病家族史 19(24.68) 9(11.84) 4.213 0.040 AHI/(次/h) 31.98±22.22 25.45±17.87 -2.004 0.047 ODI/(次/h) 29.28±23.49 22.46±16.92 -2.063 0.041 MSpO2/% 92.64±3.24 94.50±3.37 3.482 0.001 LSpO2/% 70.61±12.05 78.04±8.97 4.327 <0.001 OSAHS: 阻塞性睡眠呼吸暂停低通气综合征; BMI: 体质量指数; AHI: 呼吸暂停低通气指数; ODI: 氧减指数; MSpO2: 平均脉搏血氧饱和度; LSpO2: 最低脉搏血氧饱和度。 2.2 OSAHS患者伴2型糖尿病的影响因素分析
二元Logistic回归分析结果显示,年龄小(OR=0.247, 95%CI为0.110~0.553, P=0.001)、BMI低(OR=0.366, 95%CI为0.152~0.881, P=0.025)是OSAHS患者伴2型糖尿病的保护因素,高脂血症(OR=2.337, 95%CI为1.068~5.113, P=0.034)、LSpO2降低(OR=3.511, 95%CI为1.132~10.891, P=0.030)是OSAHS患者伴2型糖尿病的危险因素,见表 2。
表 2 OSAHS患者伴2型糖尿病影响因素的二元Logistic回归分析因素 B SE Walds P OR 95%CI 年龄/岁 -1.399 0.412 11.554 0.001 0.247 0.110~0.553 BMI/(kg/m2) -1.004 0.448 5.033 0.025 0.366 0.152~0.881 高脂血症 0.849 0.399 4.517 0.034 2.337 1.068~5.113 糖尿病家族史 0.979 0.512 3.654 0.056 2.661 0.976~7.258 AHI/(次/h) 0.259 0.667 0.151 0.697 1.296 0.351~4.787 ODI/(次/h) 0.469 0.732 0.41 0.522 1.598 0.380~6.712 MSpO2/% 0.762 0.457 2.772 0.096 2.142 0.874~5.251 LSpO2/% 1.256 0.578 4.728 0.030 3.511 1.132~10.891 OSAHS: 阻塞性睡眠呼吸暂停低通气综合征; AHI: 呼吸暂停低通气指数; ODI: 氧减指数; BMI: 体质量指数; MSpO2: 平均脉搏血氧饱和度; LSpO2: 最低脉搏血氧饱和度。 2.3 LSpO2对OSAHS患者伴2型糖尿病的预测价值
ROC曲线结果显示, LSpO2预测OSAHS患者伴2型糖尿病的曲线下面积为0.695(95%CI为0.613~0.777, P<0.001), LSpO2最佳截断值为76.50%(敏感度54.5%, 特异度75.0%), 见图 1。
3. 讨论
OSAHS是临床公认的可引起代谢紊乱尤其是2型糖尿病的独立危险因素[7]。OSAHS患者反复发生的低通气事件会导致IH, 使氧化-抗氧化系统失衡,最终引起体内炎性通路的激活[8-9]。据研究[10-11]报道,慢性炎症反应可能参与胰岛素抵抗和β细胞功能障碍的过程,增加了2型糖尿病的发生风险。缺氧和炎症之间存在密切联系,低氧可通过激活核因子κB(NF-κB)和缺氧诱导因子-1α(HIF-1α)刺激肿瘤坏死因子-α(TNF-α)、白细胞介素-1(IL-1)等各种炎症因子的产生[12]。近年来研究[13]发现, HIF-1α是一种低氧状态下在葡萄糖代谢中起重要作用的因子,可能是胰岛素抵抗和糖尿病发展的原因。SpO2降低是OSAHS患者的主要病理生理变化之一,其引起的氧化应激和炎症反应可能是OSAHS患者伴发2型糖尿病的关键因素[14]。
IH诱导的葡萄糖代谢受损的病理生理学和分子机制尚未完全清楚,但研究[15]证明,夜间IH与葡萄糖代谢受损关系密切, IH可能对OSAHS患者2型糖尿病的发生和进展起关键作用。IH对胰腺β细胞功能、胰岛素抵抗的有害影响已在动物研究中得到证实。一项动物实验[16]发现,慢性IH可能促进导致胰腺β细胞损伤和功能障碍的线粒体源性活性氧的产生。IH还可增加肝脏中磷酸烯醇式丙酮酸羧激酶、葡萄糖6-磷酸酶等糖异生酶的基因表达,引起空腹高血糖,导致2型糖尿病[17]。此外, IH通过诱导脂肪组织的炎症及脂肪分解、影响骨骼肌中葡萄糖摄取和代谢、激活交感神经产生儿茶酚胺等途径增强外周组织和细胞的胰岛素抵抗[2]。
流行病学研究[7]表明, PSG监测的SpO2参数与2型糖尿病的风险预估相关, OSAHS患者的夜间低氧血症与2型糖尿病的早期发病具有相关性,较低的LSpO2水平可使OSAHS患者发生2型糖尿病的年龄更具年轻化的趋势。日本一项前瞻性研究[18]也发现,脉搏血氧仪评估的LSpO2等参数与2型糖尿病的发生风险有关。本研究发现, OSAHS伴2型糖尿病组的LSpO2水平显著低于单纯OSAHS组,进一步分析发现, LSpO2降低是OSAHS患者伴2型糖尿病的危险因素, LSpO2水平降低对OSAHS并发2型糖尿病具有一定的预测价值,与国内外研究[19-22]结果相似。研究[19-20]发现, SpO2水平与OSAHS严重程度有关,可用于预测OSAHS合并代谢综合征的发生。相关研究[21-22]证实,低氧血症及PSG监测中氧饱和参数的降低可能与糖化血红蛋白(HbA1c)水平升高有关。本研究推测, LSpO2降低促使OSAHS患者发生2型糖尿病的原因可能是夜间低氧血症诱导的氧化应激与炎症反应,促进胰岛β细胞损伤、功能障碍及外周组织、细胞的胰岛素抵抗。
持续气道正压通气(CPAP)是OSAHS患者的一线治疗方法,可有效改善低氧状态,缓解嗜睡、觉醒、乏力等症状,提升患者的生活质量,且可降低OSAHS合并2型糖尿病患者的血糖及HbA1c水平。但目前关于纠正低氧状态在改善OSAHS合并2型糖尿病患者并发症发生情况、长期血糖控制中的作用的研究仍较缺乏,未来还需进一步探讨LSpO2干预措施对OSAHS合并2型糖尿病患者远期预后的作用。此外,本研究为单中心研究,数据来源较单一,今后仍需开展大样本量多中心临床研究及纵向研究进一步验证结论和探索具体机制。
综上所述,夜间LSpO2水平与OSAHS患者2型糖尿病的发生有关,且低水平LSpO2可能对OSAHS患者伴发2型糖尿病有一定预测价值。LSpO2便于临床监测且易被患者接受,有望成为一种理想的评估OSAHS患者伴发2型糖尿病的参考指标,而改善低氧状态、上调LSpO2水平或可成为OSAHS合并2型糖尿病患者新的治疗思路与手段。
-
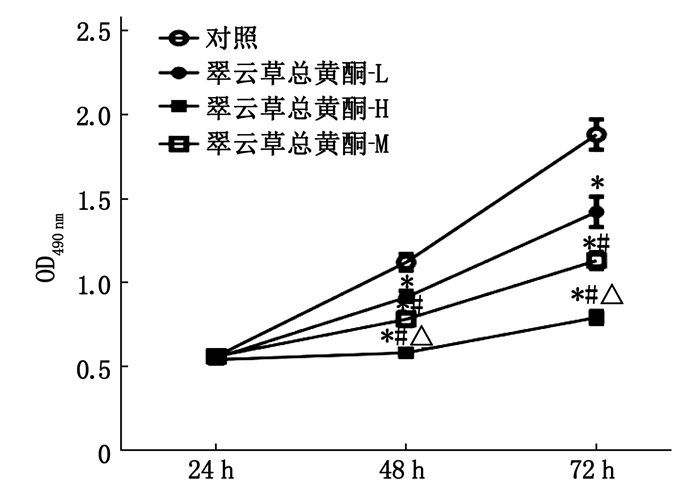
![]()
图 1 翠云草总黄酮对Caco-2细胞活性的影响
与对照组比较, *P < 0.05;
与翠云草总黄酮-L组比较, #P < 0.05;
与翠云草总黄酮-M组比较, △P < 0.05。![]()
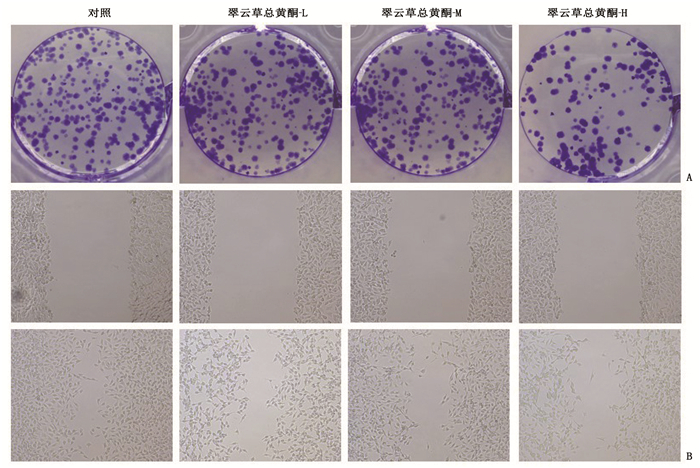
图 2 翠云草总黄酮对Caco-2细胞集落形成和迁移距离的影响
A: 克隆形成实验检测翠云草总黄酮对Caco-2细胞集落形成的影响; B: 划痕实验检测翠云草总黄酮对Caco-2细胞迁移的影响。
![]()
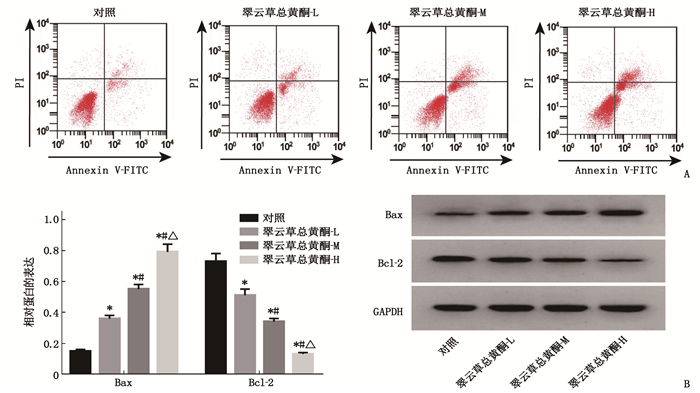
图 3 翠云草总黄酮对Caco-2细胞凋亡及Bax、Bcl-2蛋白表达的影响
A: 流式细胞术检测翠云草总黄酮对Caco-2细胞凋亡的影响; B: Western Blot检测翠云草总黄酮对Caco-2细胞中Bax和Bcl-2蛋白表达的影响。与对照组比较, *P < 0.05; 与翠云草总黄酮-L组比较, #P < 0.05; 与翠云草总黄酮-M组比较, △P < 0.05。
![]()
图 4 翠云草总黄酮对Caco-2细胞中circ_0006528和miR-330-3p表达的影响
A: qRT-PCR检测翠云草总黄酮对Caco-2细胞中circ_0006528表达的影响;
B: qRT-PCR检测翠云草总黄酮对Caco-2细胞中miR-330-3p表达的影响。
与对照组比较, *P < 0.05; 与翠云草总黄酮-L组比较, #P < 0.05; 与翠云草总黄酮-M组比较, △P < 0.05。![]()
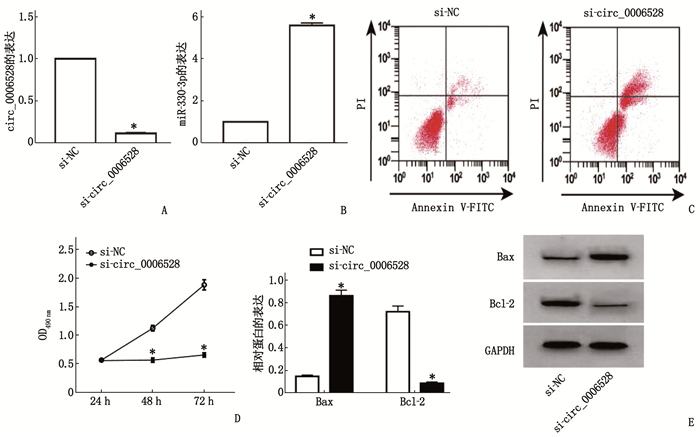
图 5 干扰circ_0006528对Caco-2细胞活性、凋亡和Bax、Bcl-2蛋白表达的影响
A: qRT-PCR实验检测转染si-circ_0006528的Caco-2细胞中circ_0006528表达; B: qRT-PCR实验检测干扰circ_0006528对Caco-2细胞中miR-330-3p表达的影响; C: 流式细胞术检测干扰circ_0006528对Caco-2细胞凋亡的影响; D: MTT实验检测干扰circ_0006528对Caco-2细胞活性的影响; E: Western Blot检测干扰circ_0006528对Caco-2细胞中Bax和Bcl-2蛋白表达的影响。与si-NC组比较, *P < 0.05。
![]()
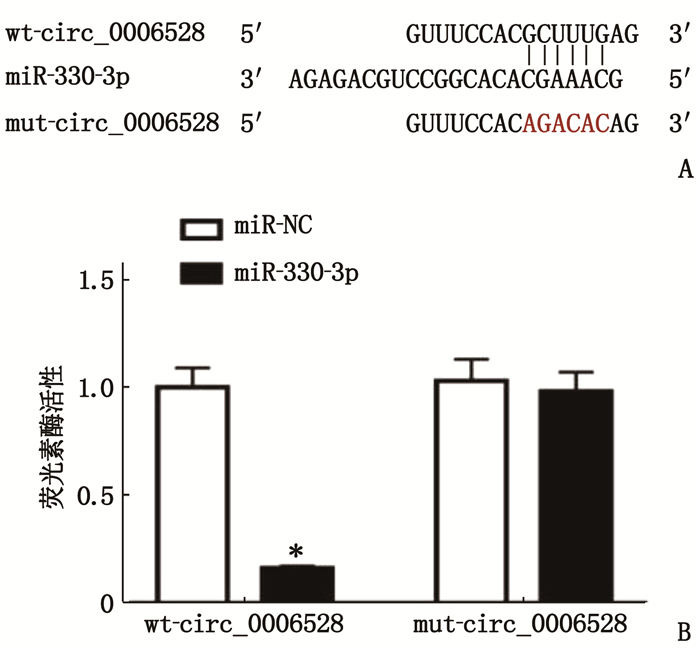
图 6 circ_0006528和miR-330-3p互补序列及荧光素酶活性检测结果
A: Circular RNA Interactome靶基因软件预测显示的circ_0006528和miR-330-3p的互补核苷酸序列;
B: 荧光素酶活性检测结果。
与miR-NC比较, *P < 0.05。![]()
图 7 过表达circ_0006528逆转翠云草总黄酮对Caco-2细胞增殖、凋亡和及Bax、Bcl-2蛋白表达的影响
A: qRT-PCR验检测翠云草总黄酮对转染pcDNA-circ_0006528的Caco-2细胞中circ_0006528表达的影响;
B: qRT-PCR检测翠云草总黄酮对转染pcDNA-circ_0006528的Caco-2细胞中miR-330-3p表达的影响;
C: 流式细胞术检测翠云草总黄酮对转染pcDNA-circ_0006528的Caco-2细胞凋亡的影响;
D: MTT实验检测翠云草总黄酮对转染pcDNA-circ_0006528的Caco-2细胞活性的影响;
E: Western Blot检测翠云草总黄酮对转染pcDNA-circ_0006528的Caco-2细胞中Bax和Bcl-2蛋白表达的影响。
与翠云草总黄酮组比较, *P < 0.05。表 1 翠云草总黄酮对Caco-2细胞集落形成和迁移距离的影响(n=3)(x±s)
组别 集落形成数/个 迁移距离/μm 对照组 115.00±5.35 166.42±8.94 翠云草总黄酮-L组 91.67±4.99* 132.25±6.44* 翠云草总黄酮-M组 71.67±2.62*# 97.89±3.24*# 翠云草总黄酮-H组 48.00±1.63*#△ 73.39±1.60*#△ 与对照组比较, *P < 0.05;
与翠云草总黄酮-L组比较, #P < 0.05;
与翠云草总黄酮-M组比较, △P < 0.05。 下载: 导出CSV
下载: 导出CSV
表 2 翠云草总黄酮诱导Caco-2细胞凋亡(n=3)(x±s)
组别 凋亡率/% Bax Bcl-2 对照组 7.40±0.31 0.15±0.01 0.73±0.05 翠云草总黄酮-L组 13.01±0.54* 0.36±0.02* 0.51±0.04* 翠云草总黄酮-M组 17.73±0.76*# 0.55±0.03*# 0.34±0.02*# 翠云草总黄酮-H组 22.42±1.04*#△ 0.79±0.05*#△ 0.13±0.01*#△ 与对照组比较, *P < 0.05; 与翠云草总黄酮-L组比较, #P < 0.05; 与翠云草总黄酮-M组比较, △P < 0.05。
下载: 导出CSV
表 3 干扰circ_0006528对Caco-2细胞集落形成、迁移和凋亡的影响(n=3)(x±s)
组别 集落形成数/个 迁移距离/μm 凋亡率/% Bax Bcl-2 si-NC组 115.00±7.48 167.04±9.48 7.84±0.67 0.15±0.01 0.72±0.05 si-circ_0006528组 41.00±0.82* 62.30±2.85* 24.46±0.85* 0.86±0.05* 0.09±0.01* 与si-NC组比较, *P < 0.05。
下载: 导出CSV
表 4 过表达circ_0006528逆转翠云草总黄酮对Caco-2细胞增殖、迁移和凋亡的影响(n=3)(x±s)
组别 circ_0006528 miR-330-3p 集落形成数/个 迁移距离/μm 凋亡率/% Bax Bcl-2 翠云草总黄酮 0.20±0.01 4.31±0.10 47.33±2.62 73.56±1.90 22.54±1.09 0.78±0.06 0.14±0.01 翠云草总黄酮+pcDNA-circ_0006528 0.83±0.06* 1.22±0.06* 103.33±3.86* 144.84±5.69* 8.63±0.54* 0.19±0.01* 0.64±0.05* 与翠云草总黄酮组比较, *P < 0.05。
下载: 导出CSV
-
[1] HAO L F, YUN Y F, LIANG R, et al. Long non-coding RNA TUSC7 suppressed colorectal cancer progression via regulation of miR-23b/PDE7A Axis[J]. Clin Invest Med, 2020, 43(4): E35-E43. doi: 10.25011/cim.v43i4.34703
[2] 邱宏聪, 刘布鸣, 陈小刚. 翠云草的研究进展[J]. 中医药导报, 2015, 21(21): 89-92. https://www.cnki.com.cn/Article/CJFDTOTAL-HNZB201521039.htm [3] 舒姮, 毛知娟, 杨勇, 等. 翠云草总黄酮对肺癌细胞生长的抑制作用[J]. 中药与临床, 2019, 10(1): 27-29. https://www.cnki.com.cn/Article/CJFDTOTAL-LCZY201901009.htm [4] CHEN L, ZHOU H, GUAN Z. CircRNA_000543 knockdown sensitizes nasopharyngeal carcinoma to irradiation by targeting miR-9/platelet-derived growth factor receptor B axis[J]. Biochem Biophys Res Commun, 2019, 512(4): 786-792. doi: 10.1016/j.bbrc.2019.03.126
[5] 陈明成, 贾洪涛. 肾透明细胞癌患者外周血中微小RNA-223的表达及临床意义[J]. 实用临床医药杂志, 2020, 24(6): 54-56, 60. https://www.cnki.com.cn/Article/CJFDTOTAL-XYZL202006015.htm [6] GAO D F, QI X W, ZHANG X F, et al. hsa_circRNA_0006528 as a competing endogenous RNA promotes human breast cancer progression by sponging miR-7-5p and activating the MAPK/ERK signaling pathway[J]. Mol Carcinog, 2019, 58(4): 554-564. doi: 10.1002/mc.22950
[7] HUANG Y G, SUN H, MA X, et al. HLA-F-AS1/miR-330-3p/PFN1 axis promotes colorectal cancer progression[J]. Life Sci, 2020, 254: 117180. doi: 10.1016/j.lfs.2019.117180
[8] 韩芳芳, 董昀, 朱渊, 等. 黄柏红花散对抗肿瘤药物相关静脉炎的预防效果[J]. 实用临床医药杂志, 2021, 25(13): 102-105. doi: 10.7619/jcmp.20211669 [9] 唐末, 杨宇飞, 宋卓, 等. 基于网络药理学探讨四君子汤治疗结直肠癌靶点及通路研究[J]. 国际中医中药杂志, 2022, 44(2): 206-211. doi: 10.3760/cma.j.cn115398-20200803-00017 [10] 程小伟, 吴锐荣, 张振勇, 等. 中药熏洗治疗结直肠癌患者化疗后手足综合征的疗效观察[J]. 实用临床医药杂志, 2020, 24(21): 32-35. doi: 10.7619/jcmp.202021010 [11] 叶丽芳, 高卓维. 龙葵正丁醇提取物抗人结直肠癌SW480细胞增殖及其作用机制研究[J]. 世界中西医结合杂志, 2019, 14(3): 356-358, 363. https://www.cnki.com.cn/Article/CJFDTOTAL-SJZX201903018.htm [12] 钟妙文, 叶慧青, 黎群足, 等. 芪附龙葵汤联合化疗对转移性结直肠癌患者生存质量的影响[J]. 现代中西医结合杂志, 2016, 25(18): 1980-1982. doi: 10.3969/j.issn.1008-8849.2016.18.014 [13] 孙鹏达, 孙冬. 桂枝水提物诱导人结直肠癌SW480细胞周期进程及其凋亡机制[J]. 中国老年学杂志, 2020, 40(20): 4396-4399. doi: 10.3969/j.issn.1005-9202.2020.20.042 [14] 陈兰英, 周朦静, 崔亚茹, 等. 白头翁皂苷干预糖酵解途径抑制SW480人结直肠癌细胞增殖作用研究[J]. 中药材, 2019, 42(3): 652-656. https://www.cnki.com.cn/Article/CJFDTOTAL-ZYCA201903038.htm [15] ZHANG C C, LI Y H, FENG X Z, et al. Circular RNA circ_0001287 inhibits the proliferation, metastasis, and radiosensitivity of non-small cell lung cancer cells by sponging microRNA miR-21 and up-regulating phosphatase and tensin homolog expression[J]. Bioengineered, 2021, 12(1): 414-425. doi: 10.1080/21655979.2021.1872191
[16] LI M, CHI C, ZHOU L Q, et al. Circular PVT1 regulates cell proliferation and invasion via miR-149-5p/FOXM1 axis in ovarian cancer[J]. J Cancer, 2021, 12(2): 611-621. doi: 10.7150/jca.52234
[17] LIU J Q, XUE N N, GUO Y X, et al. CircRNA_100367 regulated the radiation sensitivity of esophageal squamous cell carcinomas through miR-217/Wnt3 pathway[J]. Aging (Albany NY), 2019, 11(24): 12412-12427. http://www.ncbi.nlm.nih.gov/pubmed/31851619
[18] LIU G Q, ZHANG Z X, SONG Q, et al. Circ_0006528 contributes to paclitaxel resistance of breast cancer cells by regulating miR-1299/CDK8 axis[J]. Onco Targets Ther, 2020, 13: 9497-9511. doi: 10.2147/OTT.S252886
[19] HU C X, FANG K, ZHANG X F, et al. Dyregulation of the lncRNA TPT1-AS1 positively regulates QKI expression and predicts a poor prognosis for patients with breast cancer[J]. Pathol Res Pract, 2020, 216(11): 153216. doi: 10.1016/j.prp.2020.153216
[20] WANG H B, LIU G J, LI T, et al. miR-330-3p functions as a tumor suppressor that regulates glioma cell proliferation and migration by targeting CELF1[J]. Arch Med Sci, 2020, 16(5): 1166-1175. doi: 10.5114/aoms.2020.95027
[21] WANG Z H, QU H J, GONG W J, et al. Up-regulation and tumor-promoting role of SPHK1 were attenuated by miR-330-3p in gastric cancer[J]. IUBMB Life, 2018, 70(11): 1164-1176. doi: 10.1002/iub.1934
[22] WEI C H, ZHANG R G, CAI Q, et al. microRNA-330-3p promotes brain metastasis and epithelial-mesenchymal transition via GRIA3 in non-small cell lung cancer[J]. Aging (Albany NY), 2019, 11(17): 6734-6761. http://www.ncbi.nlm.nih.gov/pubmed/31498117/
[23] ZHAO X, CHEN G Q, CAO G M. Abnormal expression and mechanism of miR-330-3p/BTG1 axis in hepatocellular carcinoma[J]. Eur Rev Med Pharmacol Sci, 2019, 23(16): 6888-6898. http://www.ncbi.nlm.nih.gov/pubmed/31486488
[24] XIONG X C, SHI Q, YANG X J, et al. LINC00052 functions as a tumor suppressor through negatively modulating miR-330-3p in pancreatic cancer[J]. J Cell Physiol, 2019, 234(9): 15619-15626. doi: 10.1002/jcp.28209
[25] 黄开禹, 穆磊, 王向阳, 等. 结直肠癌中miR-330-3p-RUVBL1信号轴对肿瘤细胞增殖、凋亡的影响[J]. 华中科技大学学报: 医学版, 2020, 49(5): 517-523. doi: 10.3870/j.issn.1672-0741.2020.05.002



计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
 苏公网安备 32100302010246号
苏公网安备 32100302010246号