
Advances for ferroptosis in treating myocardial ischemia reperfusion injury
-
摘要:
铁死亡是一种铁依赖性的脂质过氧化介导细胞膜损伤致细胞死亡的新方式, 其受多种细胞代谢途径的调控, 包括铁代谢、脂质代谢、氧化还原系统等, 许多器官的损伤和退行性病变均与其相关, 在治疗缺血性疾病和脂质过氧化相关的退行性变疾病中有巨大潜力。心肌缺血再灌注损伤(MIRI)是急性心肌梗死患者进行血运重建治疗后最常见的死亡原因, 近年的研究表明铁死亡与MIRI密切相关, 通过氧化应激、铁代谢、脂质代谢、内质网应激、炎症反应等影响MIRI, 干预再灌注过程中的铁死亡可以有效改善心功能, 减少梗死面积。本文就铁死亡在MIRI中的具体作用及相关研究进展予以综述。
Abstract:Ferroptosis, a new form of programmed cell death marked by iron-dependent phospholipid peroxidation, is regulated by complex cellular metabolic pathways, including iron metabolism, lipid metabolism, and oxidation-reduction system, is associated with many organ injuries and degeneration, and has great potential in the treatment of ischemic diseases and lipid peroxide-related degenerative diseases. Myocardial ischemia reperfusion injury (MIRI) is the most common cause of death in patients with acute myocardial infarction after revascularization therapy. Recent studies have shown that ferroptosis is intimately related to the pathological process of MIRI. Ferroptosis is associated with MIRI through oxidative stress, iron metabolism, lipid metabolism, endoplasmic reticulum stress and inflammatory response. Intervention of ferroptosis during reperfusion can effectively improve cardiac function and reduce myocardial infarct size. In this paper, the research progress was explored between ferroptosis and MIRI, and the specific role of ferroptosis in MIRI was discussed.
-
食管癌(EC)是一种上消化道恶性肿瘤,临床病理类型主要是鳞癌和腺癌[1]。EC主要依赖手术治疗,手术前后采用新辅助化疗或同步放化疗巩固疗效[2]。局部复发和癌细胞转移是食管癌患者手术后治疗失败的重要原因。因此, EC的早期诊断和干预尤为重要,寻找有效的EC诊断指标具有重要意义。FOXO1是FOXO家族成员之一,能调节细胞凋亡、增殖、能量代谢、氧化应激反应,其突变与癌症形成密切相关[3]。SMAD4是转化生长因子(TGF)-β效应因子,其缺失与组织病理学转变、转移性疾病和预后不良相关[4]。SMAD4能影响正常的TGF-β信号传导,调节靶基因表达,在细胞生长和发育中具有重要作用[5]。研究[6]表明,微小RNA(miR)-183能靶向调节SMAD4, 从而参与食管鳞状细胞癌的侵袭和迁移。miR-183还可通过靶向调节FOXO1, 增强顺铂诱导的EC细胞凋亡途径[7]。但目前关于FOXO1和SMAD4表达与食管癌患者预后关系的研究较少。基于此,本研究探讨EC组织中FOXO1和SMAD4的表达水平,以及其与临床病理特征和预后的关系,以期为EC预后的早期诊断提供参考。
1. 资料与方法
1.1 一般资料
选取2019年1月—2020年4月本院行手术治疗的EC患者131例为研究对象,收集其手术切除的癌组织和癌旁组织标本。收集患者肿瘤部位、肿瘤直径、肿瘤分化程度、淋巴结转移等基本临床资料。131例EC患者年龄35~71岁,平均(52.86±10.37)岁; 男73例,女58例; 食管鳞状细胞癌75例,食管腺癌56例; TNM分期为Ⅱ期48例,Ⅲ期47例,Ⅳ期36例。纳入标准: ①符合EC相关诊断标准[8]且经病理学确诊者; ②首次进行EC根治术者; ③入组前未接受放疗或化疗者; ④对研究内容知情同意,且临床资料完整者; ⑤术后均进行辅助放化疗者。排除标准: ①患有免疫系统疾病者; ②患有肝、肾等重要脏器疾病者; ③患有其他恶性肿瘤或具有肿瘤家族史者; ④存在远处转移者。本研究已获得河北省退役军人总医院医学伦理委员会审核批准(批号: 2018-1038)。
1.2 方法
1.2.1 苏木精-伊红(HE)染色
采取EC组织条后,另取距EC边缘5 cm处组织(癌旁组织),制备病理蜡块,并切制4 mm厚度的组织白片。组织切片脱蜡水洗,苏木精、伊红依次染色,脱水,封片后显微镜下观察。
1.2.2 实时荧光定量聚合酶链反应(qRT-PCR)测定FOXO1和SMAD4的mRNA表达
Trizol试剂提取EC组织及癌旁组织总RNA, 反转录为cDNA, 进行PCR。以β-actin为内参,采用2-△△Ct法计算FOXO1和SMAD4的mRNA表达。引物序列见表 1。
表 1 qRT-PCR引物序列基因名称 正向引物5′-3′ 反向引物5′-3′ FOXO1 TTGGATTCCTGCCTGCTGAT CGCTTCTTGGGTTGATCTAG SMAD4 GACGGAGACGCTGGTGATAGCGTC CGGCAGTTATACCGCCAAGTCAACG β-actin AGCCATGTACGTAGCCATCC TGTCCAGCTGGTCCTTCT 1.2.3 免疫组化法测定FOXO1和SMAD4的蛋白表达
组织切片复水后,进行抗原修复后,采用免疫组化检测试剂盒对FOXO1和SMAD4蛋白表达进行测定,实验操作步骤严格按照试剂盒说明书进行。采用已知阳性组织切片作为阳性对照,采用磷酸缓冲液作为阴性对照。FOXO1、SMAD4均位于细胞核、细胞质中。结果判定: 染色程度,无色(0分)、淡黄色(1分)、棕黄色(2分)、褐色或黑色(3分); 染色细胞比例, 1分(染色细胞比例≤10%)、2分(染色细胞比例>10%~50%)、3分(染色细胞比例>50%~75%)、4分(染色细胞比例>75%); 染色程度评分与染色细胞比例评分乘积为最终得分, < 3分为阴性, ≥3分为阳性[9]。
1.2.4 随访
以电话或门诊复查方式对131例EC患者进行为期3年的随访。患者发生癌症相关死亡时,随访终止。
1.3 统计学分析
采用SPSS 25.0软件分析数据。计量资料均符合正态分布,以均数±标准差(x±s)表示,组间比较采用独立样本t检验; 计数资料以[n(%)]表示,组间比较采用χ2检验。采用Spearman相关性分析法分析EC组织中FOXO1与SMAD4表达的相关性; 采用Cox回归法分析EC患者预后的影响因素; 采用Kaplan-Meier法进行EC患者生存分析。P < 0.05为差异有统计学意义。
2. 结果
2.1 EC组织HE染色情况
EC组织形成癌巢,细胞排列紊乱,形态不一,核染色较深,存在核分裂,见图 1。
2.2 EC组织及癌旁组织中FOXO1和SMAD4表达比较
EC组织及癌旁组织中FOXO1和SMAD4蛋白表达结果见图 2。EC组织中的FOXO1 mRNA表达及其蛋白阳性表达率高于癌旁组织, SMAD4 mRNA表达及其蛋白阳性表达率低于癌旁组织,差异有统计学意义(P < 0.05), 见表 2、表 3。
![]() 图 2 FOXO1和SMAD4蛋白在EC组织及癌旁组织中表达(放大200倍)A: EC组织中FOXO1蛋白; B: 癌旁组织中FOXO1蛋白; C: EC组织中SMAD4蛋白; D: 癌旁组织中SMAD4蛋白。表 2 EC组织及癌旁组织中FOXO1和SMAD4 mRNA表达水平(x±s)
图 2 FOXO1和SMAD4蛋白在EC组织及癌旁组织中表达(放大200倍)A: EC组织中FOXO1蛋白; B: 癌旁组织中FOXO1蛋白; C: EC组织中SMAD4蛋白; D: 癌旁组织中SMAD4蛋白。表 2 EC组织及癌旁组织中FOXO1和SMAD4 mRNA表达水平(x±s)组织 n FOXO1 mRNA SMAD4 mRNA EC组织 131 1.63±0.37* 0.62±0.18* 癌旁组织 131 1.02±0.21 1.03±0.25 与癌旁组织比较, * P < 0.05。 表 3 EC组织及癌旁组织中FOXO1和SMAD4蛋白表达比较[n(%)]组织 n FOXO1蛋白表达 SMAD4蛋白表达 阳性 阴性 阳性 阴性 EC组织 131 88(59.46)* 43(37.72)* 45(36.29)* 86(62.32)* 癌旁组织 131 60(40.54) 71(62.28) 79(63.71) 52(37.68) 与癌旁组织比较, * P < 0.05。 2.3 EC组织中FOXO1和SMAD4蛋白表达与患者临床病理特征的关系
EC组织中FOXO1和SMAD4蛋白表达与患者年龄、性别、肿瘤部位、浸润程度、体质量指数(BMI)、饮酒史、吸烟史、病理类型、神经侵犯、脉管侵犯无相关性(P>0.05); EC组织中FOXO1和SMAD4表达与肿瘤直径、肿瘤分化程度、淋巴结转移、TNM分期有相关性(P < 0.05), 见表 4。
表 4 EC组织中FOXO1和SMAD4蛋白表达与患者临床病理特征的关系[n(%)]临床病理特征 分类 FOXO1阴性表达(n=43) FOXO1阳性表达(n=88) χ2 P SMAD4阴性表达(n=86) SMAD4阳性表达(n=45) χ2 P 年龄 >50岁 20(46.51) 45(51.14) 0.247 0.619 45(52.33) 20(44.44) 0.734 0.392 ≤50岁 23(53.49) 43(48.86) 41(47.67) 25(55.56) 性别 男 22(51.16) 51(57.95) 0.540 0.462 50(58.14) 23(51.11) 0.591 0.442 女 21(48.84) 37(42.05) 36(41.86) 22(48.89) 肿瘤直径 >3 cm 16(37.21) 60(68.18) 11.376 0.001 60(69.77) 16(35.56) 14.196 < 0.001 ≤3 cm 27(62.79) 28(31.82) 26(30.23) 29(64.44) 肿瘤部位 上中段 22(51.16) 43(48.86) 0.061 0.805 40(46.51) 25(55.56) 0.967 0.326 下段 21(48.84) 45(51.14) 46(53.49) 20(44.44) 分化程度 高中分化 32(74.42) 37(42.05) 12.144 < 0.001 35(40.70) 34(75.56) 14.399 < 0.001 低分化、未分化 11(25.58) 51(57.95) 51(59.30) 11(24.44) 浸润程度 肌层及以内 25(58.14) 40(45.45) 1.859 0.173 39(45.35) 26(57.78) 1.826 0.177 肌层以外 18(41.86) 48(54.55) 47(54.65) 19(42.22) 体质量指数 ≥23 kg/m2 23(53.49) 42(47.73) 0.384 0.536 46(53.49) 19(42.22) 1.500 0.221 < 23 kg/m2 20(46.51) 46(52.27) 40(46.51) 26(57.78) 饮酒史 是 21(48.84) 48(54.55) 0.378 0.539 45(52.33) 24(53.33) 0.012 0.913 否 22(51.16) 40(45.45) 41(47.67) 21(46.67) 吸烟史 是 19(44.19) 44(50.00) 0.391 0.532 45(52.33) 18(40.00) 1.798 0.180 否 24(55.81) 44(50.00) 41(47.67) 27(60.00) 淋巴结转移 是 13(30.23) 52(59.09) 9.623 0.002 54(62.79) 11(24.44) 17.377 < 0.001 否 30(69.77) 36(40.91) 32(37.21) 34(75.56) 病理类型 鳞癌 20(46.51) 55(62.50) 3.017 0.082 47(54.65) 28(62.22) 0.692 0.406 腺癌 23(53.49) 33(37.50) 39(45.35) 17(37.78) TNM分期 Ⅱ~Ⅲ期 38(88.37) 57(64.77) 8.072 0.004 56(65.12) 39(86.67) 6.884 0.009 Ⅳ期 5(11.63) 31(35.23) 30(34.88) 6(13.33) 神经侵犯 有 18(41.86) 44(50.00) 0.768 0.381 42(48.84) 20(44.44) 0.229 0.633 无 25(58.14) 44(50.00) 44(51.16) 25(55.56) 脉管侵犯 有 19(44.19) 45(51.14) 0.558 0.455 40(46.51) 24(53.33) 0.550 0.458 无 24(55.81) 43(48.86) 46(53.49) 21(46.67) 2.4 EC组织中FOXO1和SMAD4蛋白表达的相关性分析
Spearman相关性分析表明, EC组织中FOXO1与SMAD4蛋白表达呈负相关(r=-0.419, P < 0.05), 见表 5。
表 5 EC组织中FOXO1与SMAD4表达的相关性指标 SMAD4阴性表达(n=86) SMAD4阳性表达(n=45) r P FOXO1阳性表达(n=88) 70 18 -0.419 < 0.001 FOXO1阴性表达(n=43) 16 27 2.5 EC患者预后影响因素的多因素Cox回归分析
以FOXO1(阳性表达=1, 阴性表达=0)、SMAD4(阳性表达=1, 阴性表达=0)、肿瘤直径(>3 cm=1, ≤3 cm=0)、分化程度(高中分化=1, 低分化、未分化=0)、淋巴结转移(否=0, 是=1)、TNM分期(Ⅳ期=1, Ⅱ~Ⅲ期=0)为自变量,以EC患者是否生存(死亡=1, 生存=0)为因变量,进行Cox回归分析。结果表明, FOXO1、SMAD4、肿瘤直径、TNM分期、淋巴结转移、分化程度是EC患者死亡的影响因素(P < 0.05), 见表 6。
表 6 EC患者预后影响因素的Cox回归分析影响因素 B SE Wald P HR 95%CI FOXO1 0.196 0.052 14.264 < 0.001 1.217 1.099~1.348 SMAD4 0.118 0.017 48.003 < 0.001 1.125 1.088~1.163 肿瘤直径 0.334 0.075 19.798 < 0.001 1.396 1.205~1.617 分化程度 -0.340 0.096 12.520 < 0.001 0.712 0.590~0.859 淋巴结转移 0.079 0.021 14.084 < 0.001 1.082 1.038~1.127 TNM分期 0.222 0.063 12.456 < 0.001 1.249 1.104~1.413 2.6 EC组织中FOXO1和SMAD4蛋白表达与患者预后的关系
随访3年,患者3年总生存率为84.73%(111/131)。采用Kaplan-Meier法分析EC组织中FOXO1和SMAD4表达与患者预后的关系,结果表明EC组织中FOXO1阳性表达患者3年累积生存率为80.68%(71/88), 低于FOXO1阴性表达患者的93.02%(40/43), 差异有统计学意义(P < 0.05); SMAD4阴性表达患者3年累积生存率为79.07%(68/86), 低于SMAD4阳性表达患者的95.56%(43/45), 差异有统计学意义(P < 0.05), 见图 3、图 4。
3. 讨论
EC作为一种侵袭性恶性肿瘤,其全球发病率较高,且具有高致死性特征。局部晚期食管癌患者的2年生存率仅为3%[10]。在中国人群中,食管鳞状细胞癌占主导地位,已知的危险因素包括食管炎、吸烟、饮酒及遗传因素[11]。多数EC患者会出现声音嘶哑、吞咽困难、胸骨后疼痛等症状,这些症状显著影响患者的生活质量。此外, EC早期症状隐匿或无临床症状,常导致疾病发展至中晚期才被发现,且预后较差[12]。因此,寻找影响EC患者预后标志物对提高患者生存率显得尤为重要。
FOXO特征为蛋白质的N端存在保守的DNA结合结构域。FOXO1能转录介导糖异生、糖原分解、脂肪生成等多种途径,参与代谢性疾病进展,在肿瘤发生中具有重要调节作用,是癌症治疗的可行靶标[13]。研究[14]表明, FOXO1在EC组织中阳性表达率较高,其表达与肿瘤分期相关,敲低FOXO1能抑制肿瘤细胞增殖。本研究中, EC组织中FOXO1 mRNA表达及其蛋白阳性表达率显著高于癌旁组织(P < 0.05); FOXO1蛋白表达与肿瘤直径、肿瘤分化程度、淋巴结转移、TNM分期有相关性(P < 0.05), 提示FOXO1表达可能与EC发生发展有关。研究[15]表明, EC细胞中FOXO1表达水平高于良性食管细胞,其过表达与肿瘤分期和淋巴结转移有关; 高FOXO1表达水平EC患者的生存期缩短; FOXO1、肿瘤分期是EC患者独立的预后影响因素。本研究中, Cox回归分析结果表明, FOXO1、SMAD4、肿瘤直径、分化程度、淋巴结转移、TNM分期是EC患者死亡的影响因素(P < 0.05), 提示FOXO1与EC患者预后密切相关。研究[16]表明, FOXO1在食管鳞状细胞癌组织中表达上调,FOXO1阳性表达患者的临床结局较差,其过表达提示患者预后不良。本研究中, EC患者3年总生存率为84.73%,EC组织中FOXO1阳性表达患者3年累积生存率显著低于FOXO1阴性表达患者(P < 0.05), 提示FOXO1高表达可能与EC患者预后不良有关。
SMAD4在TGF-β信号通路激活中起关键作用,被认为是抑制肿瘤的基因[17]。研究[18]表明, EC组织中SMAD4显著降低, SMAD4表达上调能逆转上调miR-574-5p对EC细胞生长的促进作用; 较高的SMAD4水平表明食管鳞状细胞癌患者的预后和临床特征更好。本研究中, EC组织中SMAD4 mRNA表达及其蛋白阳性表达率显著低于癌旁组织(P < 0.05), 提示SMAD4可能参与EC进展。SMAD4蛋白表达与肿瘤直径、肿瘤分化程度、淋巴结转移、TNM分期有相关性(P < 0.05), 提示SMAD4低表达可能促进EC恶性发展。研究[19]表明, miR-181a可能通过TGF-β/SMAD4途径诱导上皮-间充质转化,从而调节癌细胞发展; SMAD4在正常组织中表达较高; 在EC患者中,其过表达与更好的生存率相关。本研究中, Kaplan-Meier分析结果表明, SMAD4阴性表达患者3年累积生存率显著低于SMAD4阳性表达患者(P < 0.05), 提示SMAD4在EC发展过程中可能具有一定作用。此外,本研究相关性分析表明, EC组织中FOXO1与SMAD4表达呈负相关(P < 0.05), 说明FOXO1和SMAD4可能存在相互作用。研究[20]表明, SMAD4可能通过调控miR-183-96-182簇直接靶向FOXO1, 进而抑制颗粒细胞的凋亡。因此,推测SMAD4可能通过抑制FOXO1调节癌细胞分裂和糖脂代谢,进而参与EC的发展和预后。本研究仍存在一定局限: 本研究样本量相对较少, EC病例选择存在差异,可能影响实验结果; 此外,本研究未结合EC公共数据集进行分析,因此后续还需进一步深入研究。
综上所述, EC组织中FOXO1阳性表达率较高, SMAD4阳性表达率较低,二者蛋白表达与患者临床病理特征有关, FOXO1和SMAD4是EC患者死亡的影响因素,或可作为预测EC预后的重要标志物,可为EC预后的早期诊断提供参考。
-
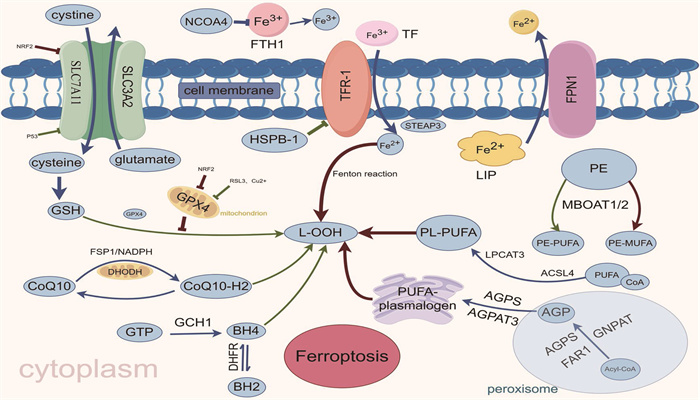
![]()
图 1 铁死亡主要调控机制
GSH: 谷胱甘肽; GSSG: 氧化型谷胱甘肽; GPX4: 谷胱甘肽过氧化物酶4; Glutamate: 谷氨酸;
SLC7A11: 溶质载体家族7成员11; SLC3A2: 溶质载体家族3成员2; Cysteine: 半胱氨酸; Cystine: 胱氨酸;
NRF2: 核因子E2相关因子2; L-OOH: 脂质过氧化物; L-OH: 脂质醇; CoQ10-H2: 还原形式的辅酶Q10;
FSP1: 铁死亡抑制蛋白1; DHODH: 二氢乳清酸脱氢酶; GCH1: 三磷酸鸟苷环化水解酶1; BH4: 四氢生物蝶呤;
BH2: 二氢生物蝶呤; GTP: 三磷酸鸟苷; DHFR: 二氢叶酸还原酶; MBOAT1/2: 膜结合O-酰基转移酶1/2;
MUFA: 单不饱和脂肪酸; PUFA: 多不饱和脂肪酸; ACSL4: 长链脂酰辅酶A合成酶4;
LPCAT3: 溶血卵磷脂酰基转移酶3; LIP: 不稳定铁池; TF: 转铁蛋白; TFR-1: 转铁蛋白受体1;
核NCOA4: 受体共激活因子4; FTH1: 铁蛋白重链1; HSPB1: 热休克蛋白β-; Acyl-CoA: 酰基辅酶A;
AGPS: 烷基甘油磷酸合成酶; FAR1: 脂肪酰基辅酶A还原酶1; GNPAT: 甘油磷酸O-酰基转移酶;
AGP: 前体1-O-烷基甘油-3-磷酸; AGPAT3: 1-酰基甘油-3-磷酸-O-酰基转移酶3;
PEDS1: 缩醛磷脂乙醇胺去饱和酶1; PUFA-plasmalogen: 多不饱和脂肪酸缩醛磷脂。 -
[1] PREM P N, SIVAKUMAR B, BOOVARAHAN S R, et al. Recent advances in potential of Fisetin in the management of myocardial ischemia-reperfusion injury-a systematic review[J]. Phytomedicine, 2022, 101: 154123. doi: 10.1016/j.phymed.2022.154123
[2] ZHAO W K, ZHOU Y, XU T T, et al. Ferroptosis: opportunities and challenges in myocardial ischemia-reperfusion injury[J]. Oxid Med Cell Longev, 2021, 2021: 9929687.
[3] YANG X, HUANG T Y, CHEN Y H, et al. Deoxynivalenol induces testicular ferroptosis by regulating the Nrf2/System Xc-/GPX4 axis[J]. Food Chem Toxicol, 2023, 175: 113730. doi: 10.1016/j.fct.2023.113730
[4] CUI Y, ZHANG Z L, ZHOU X, et al. Microglia and macrophage exhibit attenuated inflammatory response and ferroptosis resistance after RSL3 stimulation via increasing Nrf2 expression[J]. J Neuroinflammation, 2021, 18(1): 249. doi: 10.1186/s12974-021-02231-x
[5] MA S X, SUN L Y, WU W H, et al. USP22 protects against myocardial ischemia-reperfusion injury via the SIRT1-p53/SLC7A11-dependent inhibition of ferroptosis-induced cardiomyocyte death[J]. Front Physiol, 2020, 11: 551318. doi: 10.3389/fphys.2020.551318
[6] XUE Q, YAN D, CHEN X, et al. Copper-dependent autophagic degradation of GPX4 drives ferroptosis[J]. Autophagy, 2023, 19(7): 1982-1996. doi: 10.1080/15548627.2023.2165323
[7] LI W T, LIANG L, LIU S Y, et al. FSP1: a key regulator of ferroptosis[J]. Trends Mol Med, 2023, 29(9): 753-764. doi: 10.1016/j.molmed.2023.05.013
[8] MAO C, LIU X G, ZHANG Y L, et al. DHODH-mediated ferroptosis defence is a targetable vulnerability in cancer[J]. Nature, 2021, 593(7860): 586-590. doi: 10.1038/s41586-021-03539-7
[9] HU Q, WEI W H, WU D Q, et al. Blockade of GCH1/BH4 axis activates ferritinophagy to mitigate the resistance of colorectal cancer to erastin-induced ferroptosis[J]. Front Cell Dev Biol, 2022, 10: 810327. doi: 10.3389/fcell.2022.810327
[10] CHEN Y F, LI X T, WANG S Y, et al. Targeting iron metabolism and ferroptosis as novel therapeutic approaches in cardiovascular diseases[J]. Nutrients, 2023, 15(3): 591. doi: 10.3390/nu15030591
[11] TIAN H, XIONG Y H, ZHANG Y, et al. Activation of NRF2/FPN1 pathway attenuates myocardial ischemia-reperfusion injury in diabetic rats by regulating iron homeostasis and ferroptosis[J]. Cell Stress Chaperones, 2021, 27(2): 149-164.
[12] WU H, LIU Q, SHAN X Y, et al. ATM orchestrates ferritinophagy and ferroptosis by phosphorylating NCOA4[J]. Autophagy, 2023, 19(7): 2062-2077. doi: 10.1080/15548627.2023.2170960
[13] FANG Y Y, CHEN X C, TAN Q Y, et al. Inhibiting ferroptosis through disrupting the NCOA4-FTH1 interaction: a new mechanism of action[J]. ACS Cent Sci, 2021, 7(6): 980-989. doi: 10.1021/acscentsci.0c01592
[14] LIANG D G, FENG Y, ZANDKARIMI F, et al. Ferroptosis surveillance independent of GPX4 and differentially regulated by sex hormones[J]. Cell, 2023, 186(13): 2748-2764, e22. doi: 10.1016/j.cell.2023.05.003
[15] JIANG X J, STOCKWELL B R, CONRAD M. Ferroptosis: mechanisms, biology and role in disease[J]. Nat Rev Mol Cell Biol, 2021, 22(4): 266-282. doi: 10.1038/s41580-020-00324-8
[16] LIU J, KANG R, TANG D L. Signaling pathways and defense mechanisms of ferroptosis[J]. FEBS J, 2022, 289(22): 7038-7050. doi: 10.1111/febs.16059
[17] DOLL S, PRONETH B, TYURINA Y Y, et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition[J]. Nat Chem Biol, 2017, 13(1): 91-98. doi: 10.1038/nchembio.2239
[18] TANG D L, KROEMER G. Peroxisome: the new player in ferroptosis[J]. Signal Transduct Target Ther, 2020, 5(1): 273. doi: 10.1038/s41392-020-00404-3
[19] 刘丹勇, 夏正远, 韩荣辉, 等. 心肌缺血再灌注损伤机制研究的回顾与展望[J]. 中国动脉硬化杂志, 2020, 28(12): 1013-1019. https://www.cnki.com.cn/Article/CJFDTOTAL-KDYZ202012002.htm [20] RODRIGO R, GONZÁLEZ-MONTERO J, SOTOMAYOR C G. Novel combined antioxidant strategy against hypertension, acute myocardial infarction and postoperative atrial fibrillation[J]. Biomedicines, 2021, 9(6): 620. doi: 10.3390/biomedicines9060620
[21] ZINDEL J, KUBES P. DAMPs, PAMPs, and LAMPs in immunity and sterile inflammation[J]. Annu Rev Pathol, 2020, 15: 493-518. doi: 10.1146/annurev-pathmechdis-012419-032847
[22] WANG J, LIU Y, LIU Y, et al. Recent advances in nanomedicines for imaging and therapy of myocardial ischemia-reperfusion injury[J]. J Control Release, 2023, 353: 563-590. doi: 10.1016/j.jconrel.2022.11.057
[23] HEUSCH G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective[J]. Nat Rev Cardiol, 2020, 17(12): 773-789. doi: 10.1038/s41569-020-0403-y
[24] IBÁÑEZ B, HEUSCH G, OVIZE M, et al. Evolving therapies for myocardial ischemia/reperfusion injury[J]. J Am Coll Cardiol, 2015, 65(14): 1454-1471. doi: 10.1016/j.jacc.2015.02.032
[25] SPARVERO L J, TIAN H, AMOSCATO A A, et al. Direct mapping of phospholipid ferroptotic death signals in cells and tissues by gas cluster ion beam secondary ion mass spectrometry (GCIB-SIMS)[J]. Angew Chem Int Ed, 2021, 60(21): 11784-11788. doi: 10.1002/anie.202102001
[26] TANG L J, LUO X J, TU H, et al. Ferroptosis occurs in phase of reperfusion but not ischemia in rat heart following ischemia or ischemia/reperfusion[J]. Naunyn Schmiedebergs Arch Pharmacol, 2021, 394(2): 401-410. doi: 10.1007/s00210-020-01932-z
[27] YAN H F, ZOU T, TUO Q Z, et al. Ferroptosis: mechanisms and links with diseases[J]. Signal Transduct Target Ther, 2021, 6(1): 49. doi: 10.1038/s41392-020-00428-9
[28] LV Z Q, WANG F E, ZHANG X F, et al. Etomidate attenuates the ferroptosis in myocardial ischemia/reperfusion rat model via Nrf2/HO-1 pathway[J]. Shock, 2021, 56(3): 440-449. doi: 10.1097/SHK.0000000000001751
[29] KWON M Y, PARK E, LEE S J, et al. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death[J]. Oncotarget, 2015, 6(27): 24393-24403. doi: 10.18632/oncotarget.5162
[30] FAN Z Y, CAI L L, WANG S N, et al. Baicalin prevents myocardial ischemia/reperfusion injury through inhibiting ACSL4 mediated ferroptosis[J]. Front Pharmacol, 2021, 12: 628988. doi: 10.3389/fphar.2021.628988
[31] LI W Y, LI W, WANG Y, et al. Inhibition of DNMT-1 alleviates ferroptosis through NCOA4 mediated ferritinophagy during diabetes myocardial ischemia/reperfusion injury[J]. Cell Death Discov, 2021, 7(1): 267. doi: 10.1038/s41420-021-00656-0
[32] LEI D Y, LI B, ISA Z, et al. Hypoxia-elicited cardiac microvascular endothelial cell-derived exosomal miR-210-3p alleviate hypoxia/reoxygenation-induced myocardial cell injury through inhibiting transferrin receptor 1-mediated ferroptosis[J]. Tissue Cell, 2022, 79: 101956. doi: 10.1016/j.tice.2022.101956
[33] TANG L J, ZHOU Y J, XIONG X M, et al. Ubiquitin-specific protease 7 promotes ferroptosis via activation of the p53/TfR1 pathway in the rat hearts after ischemia/reperfusion[J]. Free Radic Biol Med, 2021, 162: 339-352. doi: 10.1016/j.freeradbiomed.2020.10.307
[34] LAKHAL-LITTLETON S, WOLNA M, CHUNG Y J, et al. An essential cell-autonomous role for hepcidin in cardiac iron homeostasis[J]. Elife, 2016, 5: e19804. doi: 10.7554/eLife.19804
[35] QIU M L, YAN W, LIU M M. YAP facilitates NEDD4L-mediated ubiquitination and degradation of ACSL4 to alleviate ferroptosis in myocardial ischemia-reperfusion injury[J]. Can J Cardiol, 2023, 39(11): 1712-1727. doi: 10.1016/j.cjca.2023.07.030
[36] CAI W B, LIU L, SHI X L, et al. Alox15/15-HpETE aggravates myocardial ischemia-reperfusion injury by promoting cardiomyocyte ferroptosis[J]. Circulation, 2023, 147(19): 1444-1460. doi: 10.1161/CIRCULATIONAHA.122.060257
[37] LEE Y S, LEE D H, CHOUDRY H A, et al. Ferroptosis-induced endoplasmic reticulum stress: cross-talk between ferroptosis and apoptosis[J]. Mol Cancer Res, 2018, 16(7): 1073-1076. doi: 10.1158/1541-7786.MCR-18-0055
[38] LI W Y, LI W, LENG Y, et al. Ferroptosis is involved in diabetes myocardial ischemia/reperfusion injury through endoplasmic reticulum stress[J]. DNA Cell Biol, 2020, 39(2): 210-225. doi: 10.1089/dna.2019.5097
[39] ZHOU Y Q, ZHOU H X, HUA L, et al. Verification of ferroptosis and pyroptosis and identification of PTGS2 as the hub gene in human coronary artery atherosclerosis[J]. Free Radic Biol Med, 2021, 171: 55-68. doi: 10.1016/j.freeradbiomed.2021.05.009
[40] ZHAO K, CHEN X S, BIAN Y J, et al. Broadening horizons: The role of ferroptosis in myocardial ischemia-reperfusion injury[J]. Naunyn Schmiedebergs Arch Pharmacol, 2023, 396(10): 2269-2286. doi: 10.1007/s00210-023-02506-5
[41] XU J F, ZHANG M H, LIU F, et al. Mesenchymal stem cells alleviate post-resuscitation cardiac and cerebral injuries by inhibiting cell pyroptosis and ferroptosis in a swine model of cardiac arrest[J]. Front Pharmacol, 2021, 12: 793829. doi: 10.3389/fphar.2021.793829
[42] YAN N, XU Z P, QU C H, et al. Dimethyl fumarate improves cognitive deficits in chronic cerebral hypoperfusion rats by alleviating inflammation, oxidative stress, and ferroptosis via NRF2/ARE/NF-κB signal pathway[J]. Int Immunopharmacol, 2021, 98: 107844. doi: 10.1016/j.intimp.2021.107844
 下载:
下载:




计量
- 文章访问数: 208
- HTML全文浏览量: 142
- PDF下载量: 29
 苏公网安备 32100302010246号
苏公网安备 32100302010246号